












【佳学基因检测】帕金森病、原发性神经递质疾病等多巴胺失衡系列疾病的基因矫正治疗
基因检测后怎么办?
神经系统疾病包括广泛的神经退行性疾病和神经发育疾病,这些疾病非常复杂,并且几乎普遍没有改善病情的治疗方法。因此,在为这些患者开发新的治疗策略方面,存在着巨大的未满足的临床需求。病毒基因疗法是一种很有前途的方法,其中基因传递是通过病毒载体(例如腺相关病毒和慢病毒)实现的。此类基因疗法的临床疗效已经在两种儿童期发病的神经系统疾病中得到观察到;对于脊髓性肌萎缩症和芳香族 L-氨基酸脱羧酶 (AADC) 缺乏症,基因疗法已显著改变了这些限制生命的神经系统疾病的自然病史。在这里,佳学基因检测回顾了基因治疗的最新进展,重点关注针对帕金森病和原发性神经递质疾病、AADC 缺乏症和多巴胺转运体缺乏综合征 (DTDS) 的多巴胺能基因的靶向传递。尽管最近欧洲药品管理局和药品和保健产品管理局批准了 Upstaza (eladocagene exuparvovec),这是一个重要的里程碑,但仍存在许多挑战。未来的研究需要侧重于确定临床干预的最佳治疗窗口、更好地了解治疗效果的持续时间以及改善大脑靶向性。
帕金森病、原发性神经递质疾病等多巴胺失衡系列疾病的基因矫正治疗关键词:
基因治疗、AAV、多巴胺转运蛋白缺乏综合征、AADC 缺乏症、帕金森病
介绍
用于治疗神经系统疾病的基因疗法的最新进展包括几种方法,包括基因添加、基因沉默、基因组编辑和高级编辑策略(如主要编辑)。病毒载体可以高效感染细胞,并可以传递遗传有效载荷,如互补 DNA(cDNA)、向导 RNA、小干扰 RNA(siRNA)和微小 RNA。1对于 RNA 疗法的传递,非病毒传递方法也显示出广泛应用的巨大潜力;这些应用范围从反义寡核苷酸疗法、用于治疗脊髓性肌萎缩症(SMA)的 nusinersen 到基于 mRNA 的冠状病毒病 2019(Covid19)疫苗接种。病毒载体传递的基因疗法的一个重要区别是可以通过单次给药实现终生基因治疗。然而,在儿科疾病中,尤其是转基因表达可能随着时间的推移而减弱的情况下,增强表达可能是可取的。例如,使用相同血清型的腺相关病毒 (AAV) 基因疗法的重新施用尚未在临床上实现,因为病毒载体和潜在的转基因可能会启动免疫系统。
AAV 已广泛用于中枢神经系统 (CNS) 疾病的基因治疗。单链 AAV 可以包装约 4.7 kb DNA。AAV 可以感染多种有丝分裂和有丝分裂后细胞类型。目前的共识是 AAV 不会导致人类疾病,AAV 载体被认为是无致病性的。最近有报道称,在原因不明的急性儿童肝炎病例中发现了高水平的 AAV 血清型 2。虽然这些发现并不能证明因果关系,但仍需要进一步研究以更好地了解这种关联。野生型 AAV 通过 Rep 依赖机制整合到宿主基因组中,特别是在人类 19 号染色体上的AAVS1位点,以建立潜伏期。缺乏 Rep 的AAV载体被认为是游离存在的,但在临床前 AAV 基因治疗研究中有证据表明存在随机宿主整合。病毒基因治疗中使用的重组 AAV (rAAV) 由与野生型 AAV 相同的衣壳序列和结构组成,但其基因组缺乏所有 AAV 蛋白质编码序列,而是设计了治疗性基因表达盒来代替它们。唯一来自病毒的序列是引导载体生产过程中基因组复制和包装所需的反向末端重复序列 (ITR) 。
有几种天然存在的 AAV 血清型表现出不同的组织向性,适用于不同的疾病。AAV2和 AAV9 血清型对神经元具有极好的向性,并且 AAV 附加体在转导细胞的细胞核中具有很高的耐久性。两种基于 AAV 的基因疗法已获得美国联邦药物管理局 (FDA)、欧洲药品管理局 (EMA) 和药品和保健产品管理局 (MHRA) 的批准:Luxturna(Avoretigene neparvovec‐rzyl AAV2,用于治疗视网膜色素变性)和 Zolgensma(onasemnogene abeparvovec‐xioi AAV9,用于治疗 SMA)。最近,Upstaza(eladocagene exuparvovec AAV2,用于治疗芳香族 L-氨基酸脱羧酶 [AADC] 缺乏症)已获得 EMA 和 MHRA 的批准。衣壳工程方法已被开发出来以增强特定的转导特性,从而增强肝脏或中枢神经系统的转导。合成衣壳(如 AAV PHPB 和 MNM008 )已显示出优于 AAV2 和 AAV9 的神经元转导效率,但迄今为止仅在临床前环境中进行了测试。
慢病毒载体 (LV) 主要基于逆转录病毒家族的 HIV-1,能够感染有丝分裂细胞和有丝分裂后细胞。这些载体具有更大的包装容量,可包装约 11 kb 的单链 RNA,并可长期表达转基因。10 LV临床应用的一个问题是其可能整合到宿主基因组中,尽管严格的设计修改已经缓解了这些担忧。这导致了整合缺陷型慢病毒载体的开发,这些载体在临床前研究中显示出良好的前景。
AAV 和慢病毒基因疗法均已在临床试验中针对特发性帕金森病 (PD) 进行了测试。PD 是一种渐进性神经退行性疾病,其特征是运动迟缓、表情减少、震颤、肌肉僵硬、弯腰姿势、姿势不稳定和步态蹒跚,以及许多非运动特征。它是第二大最常见的神经退行性疾病,影响着全球 600 万人。虽然运动障碍通常是患者向临床医生呈现的第一个症状,但该疾病与复杂的非运动症状有关,包括体位性低血压、便秘和抑郁症的神经精神症状、多巴胺失调症状、冲动控制受损和路易体痴呆。
帕金森病治疗的重点包括可用于管理运动症状的药物治疗策略,但没有一种能够真正改变病情。左旋多巴 ( l ‐dopa) 和其他多巴胺能药物等药物最初可能非常有益,但治疗反应会随着时间的推移而下降,在开启和关闭状态之间出现令人衰弱的波动,以及无法忍受的药物相关运动障碍。深部脑刺激 (DBS) 手术是一种选择,可能适用于对药物反应不稳定的患者,但这不是一种改变病情的治疗。靶向基因治疗的潜力使 PD 成为一种适合这种方法的疾病,因为受影响的多巴胺合成神经元特定位于黑质 (SN),尽管运动控制网络,包括边缘、中脑和脑干神经元仍然容易出现功能障碍和退化。因此,单独针对黑质纹状体系统可能只能带来部分益处。尽管 PD 中多巴胺能神经元丢失的病因尚不明确,但临床上已评估了几种基因治疗策略。这些策略主要集中在四种靶向方法上,即:(1) 恢复多巴胺合成;(2) 神经保护;(3) 基因神经调节;(4) 调节致病性葡萄糖脑苷脂酶 ( GBA ) 基因变异等疾病修饰变异。
在本综述中,佳学基因检测总结了基因治疗的进展,重点关注多巴胺能基因在 PD 和主要神经递质疾病、AADC 缺乏症和多巴胺转运体缺乏综合征 (DTDS) 中的应用。佳学基因检测还讨论了开发下一代多巴胺能疾病基因疗法的未来方向。
帕金森病的病毒基因治疗方法
帕金森病的病理特征是黑质致密部 (SNpc) 中多巴胺能神经元的逐渐丧失;这导致壳核运动区多巴胺耗竭。随着帕金森病的进展,纹状体 AADC 退化,多巴胺能 SNpc 纹状体神经末梢丧失。为了弥补帕金森病中多巴胺合成的丧失,已经开发出递送多巴胺合成酶的基因疗法。目前已有 15 项已发表的帕金森病基因治疗试验,其基因递送通过实质内立体定向注射进行。其中包括 AAV2‐GDNF(一项试验)和AAV2‐ NRTN (五项试验,其中三项关于尸检数据的出版物), AAV2 ‐GAD(四项试验),ProSavin(一项试验,一项长期随访研究)和AAV2 ‐ AADC(三项试验)。在这些临床试验中,使用统一帕金森病评定量表 (UPDRS) 报告结果 - 这是一个四部分的评估,包括临床运动检查观察(部分III ) 、运动并发症(部分IV )、影像学和通过不良事件 (AE) 报告的安全性。这些临床试验证明了帕金森病基因治疗的临床可行性和安全性。
AADC基因治疗
目前,PD 的一线药物治疗是口服左旋多巴,但随着疾病的进展,随着更多多巴胺能神经元退化和 AADC 酶(左旋多巴转化为多巴胺所需)(图 1)水平下降,临床反应也会下降。 为了解决酶水平下降的问题,已在几项 I 期开放标签研究中评估了使用 rAAV2 递送AADC的基因治疗方法。迄今为止,已有31名参与者通过双侧壳核内输注接受了 rAAV2-hAADC,以在未退化的纹状体神经元中表达 hAADC 酶 (表1)。
图 1.
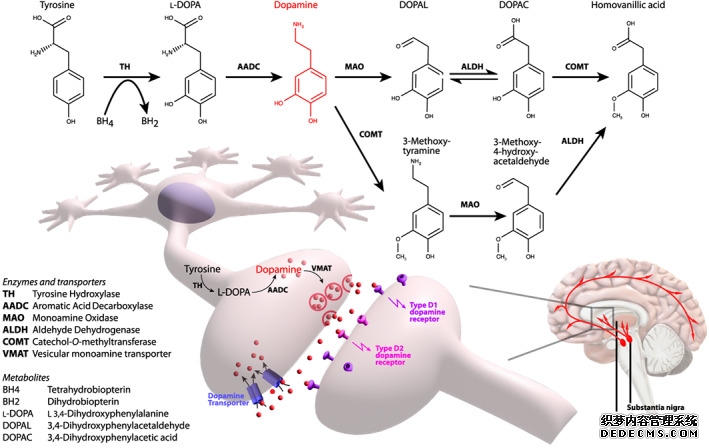
多巴胺合成和神经传递的示意图。
表 1.帕金森病多巴胺合成基因治疗临床试验
| 学习 | 剂量 (vg) 和参与者 | 患病时间(年) | 随访(月) | 结果测量 | 不良事件 |
|---|---|---|---|---|---|
|
Eberling 等人 开放标签/I 期 壳核rAAV2 |
9× 1010(200μL)n=5 | 10.8±7.5 | 6 |
28% 总 UPDRS +34% UPDRS‐III关闭状态 UPDRS开启状态、开启时间或 LED无变化 FMT‐PET 吸收率增加 30% |
手术相关无症状颅内出血 短暂性头痛和手术部位疼痛 |
|
Christine 等人 开放标签/I 期 壳核rAAV2 |
9× 1010(200μL)n=5 3×10 11(200μL)n=5 与 Eberling 等人相同的队列 9 × 10 10 |
9.3±5.3 | 6 |
31% 的总 UPDRS 和 36% 的 UPDRS-III处于非状态,32% 的 UPDRS处于工作状态 减少3.1小时休息时间 低剂量 FMT‐PET 摄取量增加 30% 高剂量 FMT‐PET 摄取量增加 75% |
2例手术相关无症状颅内出血 1 例症状性颅内出血,伴有偏瘫和失语症 |
|
Valles 等人 开放标签/I 期 壳核rAAV2 |
9× 1010(200μL)n=5 与 Eberlin Christine 等人同一队列。 |
9.3±5.3 | 6 |
壳核靶向准确性研究 结果与 Christine 等人所述一致31 |
与 Eberling 等人相同的队列。30 |
|
Muramatsu 等人 开放标签/I 期 壳核rAAV2 |
3×10 11(200μL)n=6 | 10.0±4.5 | 6 |
总 UPDRS 为 28%,UPDRS-III 为 45%(关闭) UPDRS(开启)或 LED无变化 FMT‐PET 吸收率增加 56% |
1 例静脉出血,伴有短暂性手臂无力,完全康复 |
|
Mittermeyer 等人 开放标签/I 期 壳核rAAV2 |
9× 1010(200μL)n=5 3×10 11(200μL)n=5 与 Christine 等人同一队列 |
9.3±5.3 | 48 |
长期基因表达的安全性和耐受性。 UPDRS 改善在 12 个月时恢复到手术前水平 FMT‐PET 吸收率增加 56% |
根据 Christine 等人31 4 出现短暂性运动障碍增加 |
|
Christine 等人 开放标签/I期 壳核rAAV2 |
7.5 × 10 11 (450 μL) n = 5 1.5 × 10 12 (900 μL) n = 5 4.7 × 10 12 (900 μL) n = 5 |
9.5±0.9 | 36 | F-DOPA-PET 在 6 个月时表现出剂量依赖性增加 13% ± 7%、56 ± 13%、79% ± 15%。在 12、24 和 36 个月时,UPDRS III 显著稳定或改善,LED 降低,临床和患者整体印象改善评分和 PDQ39(生活质量)降低。最低剂量组中的 1 名患者在 34 个月时进行了 DBS 插入 |
4 出现短暂性运动障碍增加 1 深静脉血栓、心房颤动和肺栓塞 1 小肠梗阻 |
|
Palfi 等人 开放标签/I/II 期 壳聚糖 左心室 TH‐AADC‐GCH |
15 1.9 × 10 7 (n = 5) 4 × 10 7 (n=5) 1 × 10 8 (n=5) |
13.9±5.3 | 12 |
UPDRS-III(关闭状态)评分提高 11.2 分,UPDRS-IV 评分提高 2 分 UPDRSII 和 PDQ‐39 的改善,LED 减少 LED 或 F‐DOPA‐PET 无显著变化 |
报告了 54 起轻度/中度事件,包括运动障碍、震颤、焦虑、开关现象 |
|
Palfi 等人 开放标签/I/II 期 壳聚糖 左心室 TH‐AADC‐GCH |
15 1.9 × 10 7 (n = 5) 4 × 10 7 (n=5) 1 × 10 8 (n=5) |
13.9±5.3 | 24–72 |
8.2 分 UPDRS-III(关闭状态)改善 其他改善未能长期维持。8 名患者在 2 年后因病情持续进展而需要 DBS 干预 |
3 严重事件 运动障碍、急性精神病和神经系统疾病(未指定) 输注后 4 年内有 1 人死亡(心肺骤停) 输注后 6 年内有 1 例死亡(腹膜炎) |
缩写:DBS,深部脑刺激;LED,左旋多巴等效剂量;PDQ39,帕金森病问卷 39;UPDRS,统一帕金森病评定量表;vg,载体基因组。
一项研究发现,6 个月时非运动状态 UPDRS-III 改善了36% , 随后在 48 个月时逐渐下降至术前基线水平。另外两项研究分别在18-39个月时观察到了类似的非运动状态 UPDRS-III 改善(改善率分别为 21%和46 %),且剂量越大,改善程度越明显。非运动状态 UPDRS-III 评分无显著变化。三项研究表明,每天的非运动状态时间增加了0.6-3.1小时,一项研究表明,无运动障碍的每天运动状态时间增加了1.5-3.3小时。 所有四项研究均进行了基因治疗后氟‐m‐酪氨酸 (FMT)‐正电子发射断层扫描 (PET) 分析,壳核摄取增加了 25%–75%,并且随着剂量的增加呈增加趋势。一项研究报告了 10 名试验参与者的 18 氟多巴 (18F-DOPA) PET 研究,显示酶活性增加了 13%–79%,并且高剂量组的增加趋势更大。五名患者出现了与手术相关的严重不良事件 (AE):三例颅内出血(两例无症状),一例静脉出血,一例深静脉血栓形成和随后的肺栓塞。八名参与者报告运动障碍暂时增加。一名受试者在输注后 34个月出现病情进展,并接受了 DBS 治疗。
2018 年,AAV2‐AADC (VY‐AADC02,NCT03562494 ) 进入 II 期试验,用于对药物反应不佳的晚期 PD 患者。该试验计划招募 85 名患者,通过枕骨途径的新递送方法递送 3.6 × 10 12 个载体基因组,并包括接受假手术的对照组。主要结果安全措施、1 年时的运动波动评估以及壳核覆盖率和酶活性。磁共振成像 (MRI) 的变化导致 FDA 在 2020 年暂停该试验的临床研究。随后,Voyager 疗法宣布其合作伙伴 Neurocrine Biosciences 终止了他们合作中的 PD 部分,从而结束了 VY‐AADC 的开发。
利用三种多巴胺合成基因进行基因治疗:GTP 环化水解酶 I、酪氨酸羟化酶和AADC
这种慢病毒基因治疗方法传递三种转基因:酪氨酸羟化酶 ( TH )、GTP 环化水解酶 I ( GCH1 ) 和芳香族 L-氨基酸脱羧酶 ( AADC ),传导纹状体神经元,旨在增加多巴胺的产生 (图 1 )。有一项已完成的临床试验涉及 15 名男性,他们通过立体定向壳核内双侧输注接受慢病毒‐TH/GCH1/AADC,这是一项 I/II 期开放标签、剂量递增研究,随后对同一队列进行了长期随访研究 (表 1 )。关闭状态的UPDRS -III 在 12 个月 (11.8 分) 和长期随访 (8.2 分)中均显着改善,剂量没有显着差异。29 在开启状态的 UPDRS-III中没有显着改善。 12 个月时 UPDRS-IV 提高了 2 分,但未能长期维持。 UPDRS -II(关闭状态:4 分;开启状态:2 分)和 PD 问卷 (PDQ)-39(提高 5.7 分)在 6 个月时有所改善,但同样未能长期维持。三名不同的患者报告了三种严重不良反应,即运动障碍、急性精神病和神经系统疾病(未指明)。所有剂量组的安全性相似,4 年和 6 年后报告了两例死亡,但被认为与治疗无关。8 名患者因病情持续进展在 2 年后需要接受 DBS 手术干预。尽管该疗法看起来很有希望,但对长期耐用性和有效性的担忧部分归因于递送和载体设计的缺陷。这促使人们开发 OXB-102,这是一种优化版本,可提高多巴胺的产生效率。随后的 AXO‐Lenti‐PD 研究 (NTC03720418) 招募了一组剂量更高、输送量和流速更高的患者,并报告称与基线相比,UPDRS-III脱靶评分显著改善,6 个月时准时性也有所改善。长期研究结果尚未报告。不幸的是,Sio Gene Therapy 于 2022 年将 Axo‐Lenti‐PD 的全球权利归还给了 Oxford Biomedica,因为他们停止了 PD 基因治疗计划。此前,Sio 首席执行官辞职,并表示资源有限。
尽管这两种针对多巴胺合成基因递送的方法颇具前景,但 PD 患者多巴胺的缺失是复杂的病理过程的结果,这些病理过程可能由遗传、表观遗传和环境等多种因素影响。因此,补充多巴胺合成机制的成分只能解决 PD 疾病过程的一个方面,可能不足以长期改善疾病。
此外,开发 PD 基因疗法还带来了其他一些科学转化挑战。控制 PD 的致病机制仍未完全了解,尽管存在转基因和化学动物模型,但没有一个能完全代表人类病理。此外,这些模型无法重现 PD 的不同阶段,人们对疾病状态和细胞病理如何影响细胞摄取和基因治疗的疗效知之甚少。随着 PD 的进展,这些基因疗法的临床疗效可能无法持续,并且将临床前研究转化为估计不同疾病阶段的有效人体剂量具有挑战性——物种之间的扩散、细胞运输以及细胞和细胞外结构存在显著差异,因此在啮齿动物模型中有效的剂量/体积可能仅覆盖人类目标的一小部分。随着时间的推移,AAV2‐AADC 在 PD 治疗中的临床试验已经增加了载体体积、输注速率和载体基因组剂量,从最初的 200 μL 中 9 × 10 10 vg 增加到最近试验中的 900 μL 中 4.7 × 10 12 vg,目的是增加壳核覆盖率。接受较高剂量的患者似乎在 3 年后表现出更持久的疗效。35有趣的是,中等剂量(900 μL 中 1.5 × 10 12)和最高剂量之间的临床疗效没有显著差异,这可能表明存在天花板效应。35可能是因为采用输注方法,较高剂量无法实现更好的转导均匀性,并且靠近注射部位的神经元被更多的病毒体转导。长期的随访研究无疑将有助于佳学基因检测了解剂量对长期临床疗效的影响。目前尚不清楚在疾病早期阶段通过治疗恢复多巴胺合成是否会比传统治疗方法具有更有意义的持续临床影响和优越性。困难在于,这类患者群体也可能是对药物治疗有反应的患者。因此,通过侵入性神经外科手术进行病毒基因治疗似乎是没有必要的,临床试验结果也未必证明其优于或比 DBS 手术更安全。在 PD 病毒基因治疗领域,证明安全性、临床疗效和优于现有治疗方法的途径一直非常具有挑战性,而且随着 Neurocrine Biosciences‐Voyager 和 Axo‐Lenti PD 项目的停止,业界对 PD 多巴胺合成基因治疗的兴趣似乎不确定。
遗传性原发性神经递质疾病的基因治疗
AADC 缺乏症
AADC 缺乏症是一种极为罕见的常染色体隐性神经发育障碍,其特征是儿茶酚胺(多巴胺、去甲肾上腺素和肾上腺素)和血清素的单胺合成受损。AADC 缺乏症患者的多巴脱羧酶 (DDC) 缺失或无功能,因此不能将左旋多巴转化为多巴胺或将 5 羟色氨酸转化为血清素(图 1)。这种疾病的原发性 AADC 酶缺陷与 PD 非常不同,PD 会逐渐出现神经退行性病变,随后 AADC 酶也会随之丧失。到目前为止,AADC 缺乏症中尚未报告神经退行性病变,且碘氟烷 I 123 DaTScan 扫描结果正常。患者有复杂的综合征,包括运动、行为、神经精神和自主神经症状。它们通常出现在婴儿期,表现为肌张力减退、眼球运动危象 (OGC) 和发育迟缓。大多数患有典型早发性疾病的患者无法控制头部,尽管也有报告称病情较轻,但少数患者可以独立行走和说话。迄今为止,文献中已描述了超过 135 名患者。39其特点是,大多数患者从标准药物治疗中获益有限 ,并且发病率高,死亡风险高。
在证明了壳核内递送 rAAV2‐AADC 基因治疗对 PD 的安全性和早期疗效后,一个合理的应用是使用该载体治疗患有遗传性 AADC 缺乏症的儿童。第一项针对 AADC 缺乏症的 rAAV2‐AADC 基因治疗研究是 2012 年的一项同情使用试验,该试验已发展到另外三项 I/II 期临床试验,将在 2022 年报告长期疗效(表 2)。壳核递送直接传导纹状体中的非多巴胺能神经元;这些包括中等棘状神经元、表达TH和 AADC 的双酶非多巴胺能神经元,以及表达 TH 并可能产生多巴胺的单酶非多巴胺能神经元。这种方法已用于三项临床试验,通过壳核内双侧输注治疗 33 名 AAV2‐AADC 儿童(n = 26,一项同情用药研究,一项 I 期开放标签,一项 I/II 期开放标签研究),随访时间为 9 至 120 个月。所有接受壳核治疗的儿童均报告临床运动改善,但继续经历 OGC。在同情试验中接受壳核基因治疗的五名儿童现已被随访 5 至 10 年。其中三人的阿尔伯塔婴儿运动评分(AIMS)和皮博迪发育运动评分-2(PDMS-2)评估保持稳定。一名儿童的膝关节生长板问题影响了评估分数,可能表明能力下降,但在接受骨科手术后,他的运动功能稳定下来。另一名患者在 5 年时出现 PDMS-2 和 AIMS 评分下降,这归因于检查引起的肌张力障碍,该障碍通过水疗法治疗。他的脑部 MRI 无异常,PET 扫描显示 AADC 表达稳定,脑脊液 (CSF) 高香草酸 (HVA) 水平稳定。报告了四起严重不良事件:一例与手术相关的无症状硬膜下出血;脑脊液漏出,需要在钻孔上放置钛网;一例危及生命的高热;一例呼吸暂停发作次数暂时增加。报告有两人死于乙型流感脑炎。
表 2.rAAV2‐AADC 临床试验,用于治疗 AADC 缺乏症
| 研究 | 參與者 | 年龄岁 | 跟进 | 结果测量 | 不良事件 |
|---|---|---|---|---|---|
|
Hwu 等人 同情用双侧壳核输注 |
4 1.6×10 11 vg‐320 μL |
4.2–6.2 | 9–24 |
安全性和耐受性 所有患者的 AIMS 和 PDMS‐2 运动评分均有改善 所有患者的 CDIIT 改善 三名患者的壳核 18F‐DOPA 摄取增加 |
所有患者均有短暂性舞蹈症运动障碍 一名患者呼吸暂停发作次数暂时增加 |
|
Chien 等人 开放标签 I 期双侧壳核输注 |
14 1.8×10 11 vg‐320 μL |
1.7–8.4 | 24 |
对运动发育和脑脊液 HVA 和 5‐HIAA 的影响 PDMS-2 分数增加(中位数:62 分) HVA CSF 浓度升高(中位数:25 nmol/L) 5‐HIAA CSF 浓度无明显变化 |
所有患者均出现短暂性运动障碍(使用利培酮可缓解) 31 例治疗相关不良事件,其中 1 例严重(危及生命的高热) 1 例因乙型流感脑炎死亡,与治疗无关 |
|
Kojima 等人 开放标签/I/II 期双侧壳核输注 |
6 2×10 11 vg‐200 μL |
4–19 | 24 |
安全性和耐受性 所有患者的 AIMS 运动评分均有改善 |
所有患者均有短暂性舞蹈症运动障碍 1例与手术相关的无症状硬膜下出血 |
|
Tai 等人 长期疗效 同情和开放标签 I/2 或 2b 期双侧壳核输注 |
26 1.8 × 10 11 vg‐320 μL(n = 21) 2.4 × 1011320 μL(n = 5) |
1.7–8.5 | 12–120个月 |
所有患者的 AIMS 和 PDMS-2 运动评分改善持续 12 个月(5 年随访) HVA CSF 浓度升高 5‐HIAA CSF 浓度无明显变化 |
术后脑脊液漏 如 Hwu和Chien 等人先前报道的 |
|
Pearson 等人 开放标签/阶段剂量递增双侧 SN VTA 输注 |
7 8.3 × 10 11 vg −160 μL(n = 3) 2.6 × 10 12 vg‐160 μL(n = 4) |
4.5–9 | 7–38 |
所有患者的 OGC 和 GMFM‐88 均得到改善 所有患者 FMT‐PET 摄取情况 所有患者的 HVA CSF 浓度均升高(6 个月时中位数为 74 nmol) 5‐HIAA CSF 浓度无明显变化 |
报告了 21 起不良事件和 10 起严重不良事件 所有患者均有短暂性舞蹈症运动障碍 短暂性烦躁加重和睡眠障碍 1例在家中猝死(归因于原发疾病) |
|
NCT02926066 II 期双侧壳核输注 |
招募 12 名患者 2022 年 1 月开始 | 2–6 | 12 |
12 个月时的疗效(脑脊液神经递质浓度和 PDMS-II 的变化) 安全性和不良事件、药代动力学、FDOPA-PET 扫描的变化。 |
尚无可用数据 |
|
NCT01395641 I/II 期双侧壳核输注 |
招募 10 名患者 2022 年 1 月开始 | 超过 2 年 | 12 |
13 个月时的疗效(脑脊液神经递质浓度和 PDMS-II 的变化) 安全性和不良事件、药代动力学、FDOPA‐PET 扫描的变化。 安全性和不良事件、药代动力学、FDOPA‐PET 扫描的变化 |
尚无可用数据 |
缩写:5-HIAA,5 羟基吲哚乙酸;AE,不良事件;AIMS,阿尔伯塔婴儿运动量表;CDIIT,婴幼儿综合发育清单;CSF,脑脊液;GMFM 88,粗大运动功能测量;HVA,高香草酸;OGC,眼球运动危象;PDMS-2,皮博迪发育运动量表。
还开发了一种治疗 AADC 缺乏症的替代方法,即通过对流增强的中脑立体定向注射将 rAAV2‐AADC 递送到黑质区和腹侧被盖区 (VTA)。这种方法旨在传导中脑并利用 rAAV2 的前向轴突运输特性将 AAV2‐hAADC 递送到纹状体黑质网络。七名年龄在 4 至 9 岁之间的儿童接受了两组 AAV2‐hAADC 的对流增强递送:1.3 × 10 11 vg(n = 3)和 4.2 × 10 11 vg(n = 4)分别实现了对黑质区和腹侧被盖区 98% 和 70% 的目标覆盖率。七名儿童中的六名在基因治疗后 3 个月内OGC 得到解决,六名儿童获得正常的头部控制,4/7 名儿童可以在 12 个月内独立坐着。18个月后,两名受试者可以在支撑下行走。38基因传递 3 个月后,脑脊液 HVA 显著增加,并持续增加。一名儿童在基因治疗后 7 个月意外猝死,死因归因于潜在疾病(与治疗无关)。在基因转移后长达 6 个月的评估中,该受试者表现出积极改善。
在不同研究中,改善最大的似乎是患有轻度至中度疾病表现的患者,并且在治疗后 12 个月内病情迅速改善。中脑和壳核给药均报告 AIMS、PDMS-2 或粗大运动功能测量-88 (GMFM-88) 有所改善。年轻参与者的改善趋势更为明显,但长期研究将更好地说明这一早期观察结果。肌张力障碍发作得到改善,OGC 的频率和/或严重程度不同程度降低,但更具体地报告了中脑给药对 OGC 的改善。40 所有接受治疗的儿童的吞咽和呼吸症状均得到改善。所有参与者的认知评估都有所改善(京都心理发展量表‐认知适应和语言‐社交性,或婴幼儿综合发展清单)。护理人员也报告了喂养、情绪、自主神经功能和睡眠方面的质的改善。壳核( FMT -PET)41和中脑、壳核和尾状核摄取(18F-DOPA PET)从基线缺失改善为双侧高信号强度(表明AADC活性恢复),最长可达基因治疗后 5 年。脑脊液神经递质分析显示多巴胺代谢物与基线相比显著增加或有增加的趋势,尽管在所有关于 6 个月脑脊液取样的研究中,其水平仍低于正常参考范围。血清素代谢物的变化各不相同,无论给药目标如何,均未显示出显着增加。所有参与者在基因治疗后数周至数月内均经历了短暂性运动障碍和舞蹈样运动,但这些都可以通过药物调整来控制。
总体而言,现在有越来越多的证据表明,基因疗法可以潜在地改善 AADC 缺乏症中的多巴胺合成、神经传递和体内平衡。如所述,这表现为核心疾病表型的改善,运动能力的提高和神经发育的进步。神经发育效应背后的机制尚不清楚,但在用 LV-hAADC 治疗的患者来源的诱导多能干细胞 (iPSC) 神经元模型中的研究表明,AADC 蛋白水平的增加和酶活性的恢复与突触素蛋白和原发性神经突分支的显着增加有关。
最近,EMA 和 MHRA 已授权通过靶向壳核递送将 eladocagene exuparvovec (Upstaza) 用于 AADC 缺乏症患者。现在评估壳核内和中脑靶向递送方法的疗效差异还为时过早。中脑靶点的体积比壳核小,但与壳核(1.8 × 10 11至 2.4 × 10 11 vg)相比,中脑递送的剂量(8.3 × 10 11至 2.6 × 10 12 vg)更高。神经元多巴胺合成和代谢受到严格调控(图 1),在多巴胺常态中,关键酶受到严格调控,以防止多巴胺和多巴氧化造成过度的氧化应激。目前尚不清楚中脑 AAV2‐AADC 递送是否能为多巴胺能神经元提供更符合生理学的 AADC 恢复,因为多巴胺能神经元中存在完整的多巴胺能酶和转运蛋白。从理论上讲,中脑递送可能降低多巴胺相关氧化应激的风险。对接受中脑和壳核基因递送治疗的患者的长期数据将说明不同脑递送方法的相对优点和优势。
最后,一个重要的未来考虑是 AADC 酶也是血清素生物合成的关键,受影响的患者表现出脑血清素缺乏。这两种基因治疗方法都没有针对血清素能神经元,而且不出所料,没有观察到对脑脊液血清素代谢物的影响。这些患者残留血清素缺乏的临床表现尚未完全了解。这无疑是未来治疗发展的一个领域,因为下一代基因疗法需要考虑针对基底神经节(壳核/中脑)和脑干的潜在双重递送方法。
双重引力
rAAV2‐AADC 基因治疗 PD 和 AADC 缺乏症的临床前和临床数据启发了佳学基因检测开发 DTDS 基因疗法的努力。这是一种极为罕见的遗传性疾病,由编码多巴胺转运蛋白 (DAT) 的 SLC6A3 的双等位基因功能丧失突变引起。 DAT调节多巴胺再摄取进入突触前神经元以终止多巴胺神经传递,因此是控制多巴胺稳态的关键转运蛋白。受影响的婴儿出现运动过度、肌张力障碍和舞蹈症,随后出现帕金森病特征,伴有运动迟缓、僵硬和震颤。他们对药物疗法或 DBS 没有反应,因此基因治疗可能是解决潜在病因的合适方法。
Illiano及其同事首次在 DAT 敲除模型中进行了概念验证基因治疗,他们使用两个 AAV 载体(衣壳血清型 10),通过立体定向注射将其递送到成年 DAT 小鼠的中脑中。为了实现对多巴胺能神经元的高特异性,第一个 AAV 在截短的大鼠 TH 启动子的控制下表达 Cre 重组酶,第二个 AAV 包含小鼠 DAT,两侧是 loxP 位点,在组成型巨细胞病毒 (CMV) 启动子的控制下。尽管结果令人鼓舞,运动表型得到改善,但使用小鼠SLC6A3 cDNA 并与 Cre 重组酶共表达限制了这种方法的临床转化潜力。
佳学基因检测旨在开发一种更直接可转化的方法。佳学基因检测使用 DTDS 患者来源的中脑多巴胺能模型生成了初步的原理证明。患者来源的神经元表现出 DAT 活性受损和疾病特异性凋亡性神经变性。为了评估体外基因治疗,佳学基因检测生成了 LV-hSLC6A3 构建体,用于在第 24 天分化时进行转导,并在衍生成熟期的第 65 天进行分析。慢病毒基因治疗可恢复 DA 摄取并恢复神经元存活,且无神经变性迹象。然后,佳学基因检测使用 DAT 敲除小鼠模型将病毒 hSLC6A3 基因治疗推进到临床应用。由于针对性立体定向注射在新生小鼠中不可行,因此进行了一项使用 DAT 敲除的 rAAV9-hSyn.hSLC6A3 的新生儿基因治疗研究。 P0 幼崽接受了脑室内 rAAV9‐hSLC6A3 基因治疗,该基因治疗可传导整个大脑。这挽救了 DAT 敲除小鼠 (DAT KO) 模型,使其恢复正常生存和运动行为,但前额皮质中的脱靶表达显示出神经元丢失和星形胶质细胞炎症反应。为了将表达限制在 DAT 表达的目标 SN 和 VTA,佳学基因检测随后通过立体定向中脑注射递送了 rAAV2‐SLC6A3,在两个对数剂量范围研究中直接模拟未来的临床应用。在这里,KO 模型中的生存和运动活动得到了完全挽救。中脑中 DAT 表达持续,AAV2‐SLC6A3 的顺向运输导致高剂量治疗动物的纹状体表达,但没有神经病理学。佳学基因检测的数据表明,立体定向中脑递送 rAAV2‐SLC6A3 基因治疗可在 DTDS 小鼠模型中以明显的安全性纠正 DAT 功能,从而加速其临床应用。
未来可能的发展
将基因疗法转化为临床疗效是一个反复的过程。13 年来,rAAV2‐AADC 载体已在 10 项 PD 和 AADC 缺乏症临床试验中进行了研究,并不断改进输送方法、输注量和速率、图像引导和载体剂量。有趣的是,载体设计没有进行任何修改。
多巴胺能疾病的基因治疗涉及复杂的挑战,需要针对特定的大脑区域和细胞类型。病毒载体技术在提高多巴胺能选择性方面的进展主要集中在 AAV 衣壳和启动子的开发上。方法越来越复杂,可能会克服当前遇到的一些障碍。一个关键的挑战是在目标细胞类型中实现生理水平的转基因表达。为了推进基因治疗,佳学基因检测必须了解基因和蛋白质表达水平在正常神经发育过程中如何变化,因为内源性水平可能在不同的发育时间点有所不同。这对于定制基因疗法以在发育儿童的一生中持续发挥作用可能至关重要。此外,基因表达水平可能因疾病阶段而异,因此需要开发对此作出反应的基因疗法,例如,解决 PD 进行性神经退行性疾病中 AADC 酶或神经营养因子水平下降的问题。
已经开发出两种较新的临床前 rAAV-AADC 基因疗法,可提供 SN、VTA 和背缝中更广泛的神经元转导。在早期的一项研究中,P0 新生 AADC 敲入 Ddc KI小鼠接受双侧脑室注射(每只幼崽总共 4 × 10 10 vg)rAAV9.CMV.hAADC。52经治疗的小鼠体重较高,存活率为 90%,而未经治疗的 Ddc KI小鼠的存活率为65% 。基因治疗改善了后肢紧握和心血管异常。检测到的 AADC 酶水平超生理水平是野生型的两倍。多巴胺水平显著升高(相当于野生型),而血清素和 3-O-甲基多巴有所改善,但未恢复到野生型水平。经治疗的 Ddc KI小鼠更加活跃,并且观察到 AADC 在整个大脑的神经元和星形胶质细胞中广泛表达。尽管证明了治疗效果,但脱靶表达可能导致异位多巴胺合成,这可能是行为过度活跃的原因。为了改善选择性神经元表达,评估了含有突触蛋白启动子的酪氨酸突变假型 AAV9/3 载体,该载体表达带有 WPRE 的小鼠 AADC(AAVN-AADC)。在本研究中,与未经治疗的 Ddc KI和野生型小鼠相比,P7 Ddc KI通过腹膜内注射 4.6 × 10 11 vg AAVN-mAADC 或 AAV9-CMV-hAADC接受全身基因治疗。从 P21 开始,两个治疗组的体重均有所增加,存活率分别提高了 95% 和 78%。系统性基因治疗挽救了行为表型,AAVN-mAADC 可使大脑多巴胺和血清素水平升高。AAV9-hAADC 观察到的过度活跃在 AAVN-mAADC 治疗的 Ddc KI中没有出现。53该载体导致 AADC 的选择性神经元表达,但腹膜内递送导致脱靶肝脏表达(尽管在小鼠中未观察到肝毒性或免疫反应)。这些研究提供了重要的概念验证:(1) AADC 在大脑中的更广泛表达不仅与多巴胺水平升高有关,还与脑血清素升高有关;(2) 使用非靶向递送方法观察到的脱靶效应会带来安全问题,需要避免。如前所述,应考虑转导血清素能背缝核,但它是脑干内一个极具挑战性的神经外科靶点。这项临床前研究展示了使用新型衣壳和神经元选择性启动子来改善靶向表达和安全性的概念验证方法。PD 临床试验和 Upstaza eladocagene exuparvovec 中使用的 AAV2‐AADC 载体受普遍存在启动子 (CMV) 的转录控制。目前的 AAV 载体不会针对特定细胞类型进行选择性表达,佳学基因检测已经开始使用神经元启动子和 AAV2 衣壳立体定向递送来改善靶向表达。目前正在进行小细胞类型特异性启动子的临床前开发,这些启动子可能通过限制在疾病靶细胞中的表达来提高表达效率和安全性。
过去 20 至 30 年来,AAV 衣壳工程主要以两种方式发展:定向进化和合理设计。在定向进化系统中,对已知血清型应用随机改组衣壳基因的过程(例如将肽插入 AAV 衣壳或噬菌体展示的已知位点)。合理设计以改进衣壳结构以实现所需特性(例如破坏天然细胞结合基序并在 Cap 基因中插入高亲和力配体)是一种替代策略。有趣的是,在临床前应用中,很少有合成衣壳取代野生型变体,也没有一种进入临床试验,因为改进的新特征很少从宿主物种(最常见的是最初在小鼠中筛选)转移到非人类灵长类动物或人类。已经进行了多次努力来生成用于 PD 的合成衣壳,其对纹状体黑质通路的转导效率更高。通过使用 BRAVE 系统进行合理设计,已对 HEK293T 细胞、原代皮质神经元、移植了源自人类胚胎干细胞的 DA 神经元的大鼠以及人类 iPSC 衍生的类器官进行了向性筛选。通过这种策略,已鉴定出 25 种性能优于野生型 AAV2 的合成衣壳,包括对大鼠和人类多巴胺能神经元具有更高逆向感染性的 MNM008 衣壳。9可以想象,这种优异的 AAV 衣壳将在未来多巴胺能系统疾病基因疗法的开发中发挥重要作用。
结论
随着时间的推移,越来越多的证据表明多巴胺基因疗法对与多巴胺失衡相关的运动障碍有效。最令人信服的数据来自针对 AADC 缺乏症的 rAAV2‐AADC 基因治疗方法。针对此类单基因疾病的基因治疗方法具有产生重大影响的潜力。PD 和 AADC 缺乏症的试验表明,疾病阶段可能会影响临床结果,这强调了治疗窗口的重要性。在临床前和临床层面,多巴胺疾病基因疗法的开发面临许多挑战;这些挑战包括改善基因表达的传递、分布和控制,以及改善多巴胺疾病的临床前建模。开发下一代基因疗法的未来方向包括设计新型多巴胺特异性衣壳和启动子,以针对这些疾病定制基因疗法,改善目标患病细胞中的基因表达并最大限度地减少脱靶效应。具有临床影响的 PD 基因疗法将需要更复杂的方法来应对神经退行性疾病的复杂性。病毒介导的多巴胺酶替代策略只是众多新治疗方法中的一种。阻止或改善异常 α‐突触核蛋白折叠、改善线粒体功能、恢复多巴胺合成、改善突触功能障碍、提高神经元存活率或细胞替代疗法是未来的策略,潜在的联合方法可能迈出阻止 PD 疾病进展的第一步。从长远来看,未来的科学发现将更好地确定病因和 PD 发病机制,以改善早期诊断,确定临床治疗窗口和潜在治疗靶点,从而实现可能逆转疾病的治疗。
(责任编辑:基因检测)